相关热词搜索:
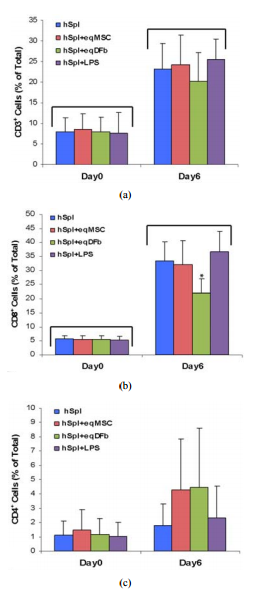
ABSTRACT The purpose of our investigationswas to measure, in a co-culture condition, the immunoresponse to allogeneic orxenogenic cells, selected as potential sources for cell therapy of arthritis.We challenged human spleen-derived cells (hSpl) by three different mechanisms:1) exposure to donor allogeneic or xenogeneic cellular antigens; 2) exposure todonor cells transduced with adenoviral antigens (Ad) and 3) lipopolysaccharide(LPS), a known inflammatory immunostimulant. The immunoresponse to allogeneichuman synovial-derived mesenchymal stromal cells alone or transduced withadenoviral green fluorescent protein (hSD-MSC or hSD-MSC/GFP) or theimmunoresponse to xenogeneic equine mesenchymal stromal cells (eqMSC) or equinedermal fibroblasts (eqDFb), characterized by the proportion of CD3+ , CD4+ ,and CD8+ human splenocytes (hSpl), was measured on Day 0 and Day 6 of co-cultureby flow cytometry. In culture with hSD-MSC, hSD-MSC/GFP, eqDFb, or eqMSC, theproportion of CD3+ and CD8+ hSpl increased with time in culture but not withexposure to cell allo- or xeno-antigens. Both hSD-MSC and hSD-MSC/GFP increasedin number during culture and were not affected in viability or proliferation byco-culture with allogeneic hSpl. In this in vitro, primary exposure study, hSpldemonstrated a natural selection and adaptation to a short-term cell cultureenvironment, and that neither allogeneic nor xenogeneic cell antigens incited agreater cellular immunoactivation than co-cultured hSpl alone. Keywords:Arthritis; Stem Cells; Allogeneic; Xenogeneic; Immunoresponse 1. IntroductionCutting-edge advancements in regenerative medicine may harness the potential ofan engineered cell source as a therapeutic vector for the repair andrestoration of multiple human tissues including articular cartilage damaged byinjury or degenerated through either osteoarthritis or rheumatoid arthritis.Allogeneic cells, from a different organism of the same species or xenogeniccells, from a different species, could provide a nearly limitless supply oftherapeutic cells for use in tissue repair. Allogeneic or xenogeneic cellsources, engineered to serve as a clinical tool, could dramatically andirrevocably enhance current medical practices that promote healing. Allogeneicmesenchymal stem cells in Phase III clinical trials, have been used as atreatment for inflammatory conditions including acute Bone Versus Graft Diseaseand Crohn’s disease [1]. In orthopaedic medicine, autologus cell-basedstrategies for tissue repair and restoration, including autologous chondroctyeimplantation and osteochondral grafts, are currently used in clinical practicewith some promising results [2-6]. A potentially promising alternative oradjunct strategy to current regenerative techniques could use allogeneic cellsor xenogenic cells. To this end, a cell-based allogeneic therapy has beenprovided for 1st generation, commercially-available cartilage neograft (DeNovo®ET Live Chondral Engineered Tissue Graft, Zimmer Holdings, Inc). This product,approved last year, relies on allogeneic, juvenile chondrocytes and aproprietary cell-scaffold system to promote healing. Potentially, allogeneic orxenogeneic cells could serve as effective therapeutic vectors in vivo tointegrate into the native biological environment of tissue. The immunoresponseof allogeneic and xenogeneic cells needs to be further investigated, andfurther observations of the cellular mechanisms and interactions will help toelucidate and describe the immunoresponse for future clinical trials. New cellsources could improve the repair and restoration of articular cartilage, suchas in injured cartilage or deteriorating cartilage as in osteoarthritis (OA),the most common form of arthritis. It is believed that there are immuneprocesses that could be responsible for the deCopyright © 2012 SciRes. OJCB 2S. S. JUMP ET AL. generation of cartilage. In particular, activated immune cellinfiltration, including T-cells, has been associated with the advancement ofarthritis [7-9]. Finding a potential cell- or tissue-based treatment fordamaged cartilage is dependent upon further understanding the immunoresponse ofcells from an articular joint, including synovial lining cells or chondrocytes.Organ or cell transplantation is characterized by an activation of hostdefenses including the activation and proliferation of immune cell typesincluding cluster determination 3 (CD3+ ) mature lymphocytes, clusterdetermination 4 (CD4+ ) T-helper (TH1) cells, and cluster determination (CD8+ )natural killer/ cytotoxic T lymphocytes (CTL) [7-9]. Allogeneicsynovial-derived cells (SDSCs) have been successful in the repair offull-thickness defects of the femoral condyle in rats; however, contaminatingmacrophages provided evidence of a delayed immune reaction to thetransplantated allogeneic cells [10]. Furthermore, xenogeneic SDSCs failed torepair cartilage defects in vivo, and an enhanced immune response,characterized by detection of major histocompatability complex antigen II(MHCII) in foreign bodies found in the repair tissue [11]. In cell culture, alimited number of studies have used a mixed immune cell design to evaluate hostversus donor reactions [12-16]. Allogeneic human MSCs derived from bone marrowsuccessfully reduced CD8+ expansion in cell culture providing support for thebeneficial immunomodulation of MSCs [17], and allogeneic fibroblast-likesynoviocytes have also reduced proliferation in T-cells found in bone marrow[12]. On the other hand, allogeneic peripheral blood mononuclear cells elicitedan activation of both CD4+ and CD8+ cells [14]. Further characterization of thepotential cellular activation of allogeneic or xenogeneic cells with the hostimmune system is needed to further develop clinical tools to control or monitorthis reaction for the future of cell therapy as a treatment for joint disease.Our study will extend the findings in the literature by using another source ofhuman immune cells, splenic tissue, to investigate the potential immuneactivation of CD3+ , CD4+ , and CD8+ cell types to allogeneic or xenogeneicmesenchymal stromal cells. The purpose of our investigations was to measure, ina co-culture condition, the immunoresponse to allogeneic or xenogenic cells,selected as potential sources for cell therapy of arthritis. We challengedhuman spleen-derived cells (hSpl) by three different mechanisms: 1) exposure todonor allogeneic or xenogeneic cellular antigens; 2) exposure to donor cellstransduced with adenoviral antigens (Ad); and 3) lipopolysaccharide (LPS), aknown inflammatory immunostimulant. For the allogeneic experiment, humansynovial-derived mesenchymal stromal cells (hSC-MSC) or hSC-MSC transduced withadenovirus expressing green fluorescent protein (hSC-MSCGFP) were co-culturedwith allogeneic hSpl. In the xenogeneic experiment, equine (eq) bone-marrowderived MSCs (eqMSC) or equine dermal fibroblasts (eqDFb) were co-cultured withhSpl. Our hypothesis was that a cell-mediated challenge of either allogeneic orxenogeneic cells would stimulate the formation and development of CD3+ and CD8+hSpl in co-culture compared to unchallenged hSpl and that adenoviral challengemay further enhance this effect. Additionally, the viability of the allogeneicor xenogeneic donor cells was expected to be reduced when co-cultured withhuman host cells. 2. Methods 2.1. Human and Equine Donor Tissue Harvest andDigestion Synovial biopsies were obtained from the knee joint of orthopedicpatients undergoing anterior cruciate ligament reconstruction by an authorsurgeon (DCF). Tissue harvest was conducted in accordance with the guidelinesset by the Institutional Review Board (IRB Protocol 2009H0256) at The OhioState University and only by consent of the patient. Synovial biopsies weredigested in sterile-filtered (0.2 um) media containing collagenase type I (0.2%m/v) for 90 min (Gibco, Carlsbad, CA). Following digestion, cells were filteredthrough a cell strainer (70 μM) and washed twice in Gey’s Balanced Salt (GBSS)(Sigma-Aldrich, St. Louis). Before initial seeding, cell samples from synoviumwere exposed to trypan blue exclusion stain and cell count and cell viabilitywas determined using a hemacytometer Bone marrow aspirates were obtained fromthe sternum of adult horses immediately after euthanasia for reasons unrelatedto the immune system. A bone marrow aspiration needle (MD Tech Inc,Gainesville, FL) was inserted into a sternebral body from the ventral aspect ofthe sternum, and marrow was aspirated into a sterile, heparin-flushed (AmericanPharmaceutical Partners Inc., Schaumburg, IL), 12-mL syringe. The procedure wasrepeated until a minimum of 10 mL of bone marrow was collected. PrimaryBMD-MSCs were isolated via centrifugation of marrow specimens and cultured in amonolayer, as has been described [18]. Derived eqBMD-MSCs (eqMSC) wereconfirmed as pluripotent by culturing in controlled osteogenic, chondrogenic,and adipogenic media containing dexamethasone with ascorbate, recombinant humantransforming growth factor-I, and dex methasone with insulin and indomethacin(Gibco, Grand Island, NY) [19-21]. Dermal fibroblasts (DFb) were obtained viaskin biopsy as part of another equine study [20,22]. Full-thickness skin tissuewas harvested using a 5 mm diameter biopsy punch from the pectoral region (10 -12 punches Copyright © 2012 SciRes. OJCB S. S. JUMP ET AL. 3 per horse) fromeach of six adult horses. The dermal layer was dissected from the epidermisunder a microscope, and DFbs were isolated by type-1 collagenase digestion(GIBCO, Grand Island, NY) and cultured in DMEM supplemented with L-glutamine(300 mg/mL), penicillin (30 mg/mL), streptomycin (30 mg/mL), and 10% fetalbovine serum at 37˚C in a 5% CO2 atmosphere. Synovial-derived mesenchymalstromal cells were passaged a minimum of 4 times, but no more than 7 times.Previous studies in the literature have demonstrated that hSD-MSCs cellsmaintain a consistent phenotype between passages 3 and 8 [23,24]. Equine BMD- MSCand Dfb were low passage (<3 passages). 2.2. Tissue Harvest of Host Mixed Immune Cells Human spleens were selected as the host mixed cell population for co-culture experiments. Approval for receiving portions of human spleens was granted by Lifeline of Ohio, an organ donor center in Columbus, Ohio. Splenocytes were harvested and prepared with high yield and successfully grown in cell culture using methods adapted from murine splenocyte culture [25,26]. Briefly, tissue was trimmed into small pieces and thoroughly minced using a syringe plunger inside of a nylon cell strainer (70 μm) in a 35 mm cell culture plate. Cells were digested using Ack lysing buffer (Gibco, Carlsbad, CA), and the samples were subjected to consecutive washes in ice cold PBS. Isolated splenocytes were immediately allocated to cultures as described below in experimental design. 2.3. Adenoviral Transduction of Donor Allogeneic Synovial-Derived Mesenchymal Stromal Cells Recombinant, E1-deficient, serotype-5 adenovirus vectors containing the open reading frame segment of human GFP (AdGFP) under the control of the cytomegalovirus promoter were generated. Successful transduction of AdGFP was verified in cell culture. Viral titer [infection units per mL, (IFU/mL)] was determined (Adeno-X Rapid TiterKit; Clonetech, Mountain View, CA, USA) and SD-MSCs were transduced at 100 multiplicities of infection (MOI) or 1 × 102 infectious units per cell [19- 21]. 2.4. Cell Co-Culture Conditions and Experimental Design For the overall experimental design, three sources of donor cells, hSD-MSC (allogeneic; Experiment 1) and eqMSC and eqDFb (xenogeneic; Experiment 2), were seeded at 10,000 cells/cm2 onto 24-well cell culture plates at Day –2 in 10% FBS containing 1% penicillin streptomycin in DMEM (Invitrogen). Human SD-MSC cultures were transduced with Ad-GFP on Day –1. For Experiments 1 and 2, isolated hSpl were obtained from 3 different human spleens for each experiment (6 spleens) and cultured in triplicate alone or added to donor cultures at a 50:1 ratio or 500,000 hSpl/cm2 on Day 0. For Experiment 1, hSpl were added to one of three co-culture immune challenges at Day 0; allogeneic hSD-MSC, allogeneic hSDMSC-GFP, or lipopolysaccharide stimulation (LPS 10 μg/mL, Sigma-Aldrich, St. Louis, MO). For Experiment 2, hSpl were assigned to one of three co-culture immune challenges at Day 0; xenogeneic eqMSC, xenogeneic eqDFb, or lipopolysaccharide stimulation (LPS 10μg/mL, Sigma-Aldrich, St. Louis, MO). On Day –1, hSD-MSC wells were assigned to receive adenoviral vector transduction. Cells were transduced with AdGFP at 100 MOI for 2 h in Gey’s Balanced Salt Solution. Effective transduction methods with AdGFP have been validated in previously published work [19]. On Day 0, prior to the addition of the allogeneic hSpl cells, GFP expression was confirmed by fluorescent microscopy and recorded. In addition to co-cultures, hSpl were cultured alone. At day 0, hSpl were removed for cellular viability staining and flow cytometry analysis approximately two hours after co-cultures were established to serve as a baseline immune status. At Days 0 and 6, hSpl were harvested for cell surface marker content as measured by flow cytometry. 2.5. Flow Cytometry, Cell Numbers, and Cell Viability Flow cytometry (C-Flow Plus, Accuri, Ann Arbor, MI) was performed on hSpl subjected to extracellular staining to determine the proportion of cells positively-stained for binding of CD3, CD4, or CD8 antibodies. Human monoclonal antibodies for each immune cell subtype (FITCconjugated-anti-CD3, APC-conjugated anti-CD4, and PEconjugated anti-CD8, R&D Systems, Minneapolis, MN) were used to assess the potential formation of immune cell types. At Days 0 and 6, we investigated formation of CD3+ , CD4+ , and CD8+ cells; an unstained control and an antibody control for each respective antibody were used on each of flow cytometry analysis. Cell number and viability for both hSD-MSC and hSD-MSC/GFP were determined on Day 6. Cells were trypsinized, washed, and subjected to a trypan blue exclusion stain. Cells were counted in a hemacyometer; the number of live cells was recorded. 2.6. Statistical Analysis For co-culture experiments, cells isolated from one human joint (hSD-MSC or hSD-MSC/GFP or one animal (eqMSC or eqDFb) and an inpidual spleen (hSpl) were Copyright © 2012 SciRes. OJCB 4 S. S. JUMP ET AL. considered an n of 1. Data for quantitative outcomes of hSpl CD3+ , CD4+ , CD8+ number and proportion, as well as donor cell number and viability were presented as means ± standard error of the mean (SEM). Two factor analysis of variance (ANOVA) for Day (0 and 6) and hSpl condition (alone, hSD-MSC, hSD-MSC-GFP; or alone, eqMSC, eqDFb, LPS) was performed for hSpl number, donor cell number, and hSpl or donor cell viability. All analyses were performed using a comercially available statistical software package (Statistical Analysis Software, SAS 9.1). Statistical significance was accepted at p ><0.05 3. Results Human spleens were obtained on ice, and splenoctyes wereharvested with initial high hSpl number and viability (>95%) (Figure 1(A))and co-cultured with donor cells at Day 0 (Figure 1(B)). The proportion of gatedviable hSpl on the Day 0 of culture measurement by flow cytometry was in therange of 65% - 74% (Figures 2(a) and 2(b)). The proportion of CD+ hSpl (CD3+ ,CD8+ , plus CD4+ ) was <15% of total gated cells on Day 0 culture measurement by flow cytometry (Figures 3 and 4). Culture of hSpl alone resulted in a significant decrease in hSpl numbers and viability to an average of approximately 31% by Day 6 as measured by flow cytometry (Figures 2(a) and 2(b)). Co-culture of hSpl with allogeneic or xenogeneic donor cells did not further decrease hSpl numbers (Figures 2(a) and 2(b)). Both allogeneic and xenogeneic donor cells proliferated in cell co-culture with hSpl (Figure 5) and were of normal morphology with high efficiency of GFP expression demonstrated in hSD-MSC-GFP cells for the entire 6 days. (Figure 1(B), insert) Synovial-derived mesenchymal stromal cells were successfully transduced (hSD-MSC-GFP) 24h after transduction with AdGFP at 100 multiplicities of infection (100 MOI). (Day 0). In the allogeneic co-culture Experiment 1, the proportion of CD3+ and CD8+ hSpl was significantly greater on Day 6 (4-fold and 5-fold, respectively) regardless of culture condition as compared to Day 0. (Figures 3(a) and 3 (b)) On Day 6, hSpl cultured in LPS had a greater proportion of CD3+ hSpl than hSpl cultured alone. There was no change in the proportion of CD4+ hSpl on Day 6 versus 0 (Figure 3(c)). In the xenogeneic co-culture Experiment 2, the proportion of CD3+ and CD8+ hSpl was significantly greater on Day 6 (3-fold and 6-fold, respectively) regardless of culture condition as compared to Day 6 as in the allogeneic co-culture experiment and compared to Day 0. (Figures 4(a) and 4(b)) On Day 6, hSpl co-cultured with eqDFb had a lesser proportion of CD8+ hSpl than hSpl cultured alone. There was no change in the proportion of CD4+ Figure 1. (A) Photomicrograph (100× magnification) of a hemacytometer containing a cell dilution of human splenocytes stained with trypan blue stain after being digested from a human spleen. Viable hSpl are yellow and circular, and non-viable human splenocytes (hSpl) are blue and pyknotic. Each spleen sample that was digested had greater than 95% living cells prior to the start of the co-culture experiments (Day 0); (B) Photomicrograph (100× magnfication) of a co-culture containing hSpl (small circular cells) and human synovial-derived mesenchymal stromal cells (hSD-MSC) 100x magnification. Human SD-MSC transduced with AdGFP at 100 infectious Ad particles per cell, had >95%expression of GFP under fluorescent microscopy at 24 hours (insert; 200×magnification). cells between days (Figure 4(c)). 4. Discussion This studyprovided evidence that, in a short-term culture environment, hSpl undergo anatural selection and adaptation to cell culture conditions. In the allogeneicor xenogenic co-culture conditions, no particular evaluated cell source(allogeneic synovial or xenogeneic bonemarrow, or skin) or donor source (humanallogeneic or equine xenogeneic) promoted survival, inhibited death, orpromoted death of the lymphocyte cell fraction of hSpl. In addition, these datashowed that neither allogeneic nor Copyright © 2012 SciRes. OJCB S. S. JUMP ETAL. 5 (a) (b) Figure 2. Number of gated (viable) hSpl on Day 0 and Day 6 in theallogeneic Experiment 1 (Panel (a)) and the xenogeneic Experiment 2 (Panel(b)). Abbreviations: hSpl = human splenocytes, hSpl + hSD – MSC = humansplenoctyes cocultured with human synovial-derived mesenchymal stromal cells,hSpl + hSD – MSC/GFP = human splenocytes cocultured with human synovial-derivedmesenchymal stromal cells transduced with AdGFP, hSpl + eqMSC = humansplenoctyes co-cultured with equine mesenchymal stromal cells, hSpl + eqDFb =human splenocytes co-cultured equine dermal fibroblasts, hSpl + LPS = humansplenocytes cocultured g/mL). Data aremwithlipopolysaccharide (LPS, 10 SEM. The bracket (p±mean < 0.05) denotes astatistical difference at Day 6 compared to Day 0. xenogeneic cells elicited alymphocytic/dendritic cell selection and no donor source promotedproliferation, at least in vitro and for this short duration of primaryexposure. The short survival of immune cells in culture (Spl or peripheralblood monocytes) limit in vitro studies. However, within these limitations, theevidence suggested that allogeneic or xenogeneic cells may be immunotolerated,at least initially, in a host environment. Our study was a first to investigatethe immunoresponse of a mixed population of hSpl to different tissue sources ofboth allogeneic and xenogeneic origin cells and assessed three differingmechanisms of immune challenge, including 1) cellular antigens, 2) adenoviralantigens, and 3) lipopolysaccharide (LPS), a mediator of the inflammatoryimmune reaction. We were unable to document (a) (b) (c) Figure 3. Human Humansplenocytes (hSpl) co-cultured with allogeneic synovial-derived mesenchymalstromal cells (hSDMSC) and subjected to extracelluar staining using antibodiesspecific for CD3, CD4, or CD8. The proportion of positive cells for eachantibody was determined by flow cytometry. (a) CD3+ hSpl on Day 0 and Day 6.(b) CD8+ hSpl on Day 0 and Day 6. (c) CD4+ hSpl on Day 0 and Day 6.Abbreviations: hSpl = human splenocytes, hSpl + hSD – MSC = human splenoctyesco-cultured with human synovial-derived mesenchymal stromal cells, hSpl + hSD –MSC/ GFP = human splenocytes co-cultured with human synovial-derivedmesenchymal stromal cells transduced with AdGFP, hSpl + LPS = human splenocytesco-cultured with SEM.±g/mL). Data are mean mlipopolysaccharide(LPS, 10 The bracket (p < 0.05)denotes a statistical difference at Day 6 compared to Day 0. # denotes anunexplained difference compared to other groups at Day 0. * denotes that hSpl +LPS at Day 6 is significantly greater than Day 0. significant activation of CD+immune cells or death of immune cells or death of allogeneic or xenogeneicdonor Copyright © 2012 SciRes. OJCB 6 S. S. JUMP ET AL. (a) (b) (c) Figure 4.Human splenocytes (hSpl) co-cultured with xenogeneic equine bone-marrow derivedmesenchymal stromal cells (eqMSC) or dermal fibroblasts (eqDFb) and subjectedto extracelluar staining using antibodies specific for CD3, CD4, or CD8. Theproportion of positive cells for each antibody was determined by flowcytometry. (a) CD3+ hSpl on Day 0 and Day 6. (b) CD8+ hSpl on Day 0 and Day 6.(c) CD4+ hSpl on Day 0 and Day 6. Abbreviations: hSpl = human splenocytes, hSpl+ eqMSC = human splenoctyes cocultured with equine mesenchymal stromal cells,hSpl + eqDFb = human splenocytes co-cultured equine dermal fibroblasts, hSpl +LPS = human splenocytes co-cultured ±g/mL).Data are mean mwith lipopolysaccharide (LPS, 10 SEM. The bracket (p < 0.05) denotes astatistical difference at Day 6 compared to Day 0. *denotes that hSpl + EqDFbwas significantly lower than other groups at Day 6. cells. Dermal fibroblastshave not been stated to be immunoprivileged as has been claimed for thesynovial fibroblast or bone-marrow derived mesenchymal proFigure 5. Cell numberof human synovial-derived mesenchymal stromal cells (hSD-MSC) and humansynovial-derived mesenchymal stromal cells transduced with adenoviruscontaining green fluorescent protein (hSD – MSC/GFP) on Day 0 and Day 6 asdetermined by hemacytometer live cell count following a trypan blue exclusionstaining Data SEM. The bracket (p±are mean < 0.05) denotes astatistical difference at Day 6 compared to Day 0. genitor and stromal cells,but our data suggested that xenogeneic dermal fibroblasts may beimmunosuppressive to CD8+ cells and would warrant further investigation.Importantly, further experiments in vivo would be necessary to more criticallyassess immunotolerance, to include longer exposure to antigens, and evaluate asecond exposure to elucidate the cellular contributions and anamnestic responseto isolated allogeneic or xenogeneic cells. To our knowledge, our splenocyteco-culture system is novel and contributed initial information on tolerance ofthese mixed immune cells to donor cells on short term exposure and supportsprevious publications claiming allogeneic juvenile chondroctyes exhibitimmunotolerance [27]. Previous work has also provided evidence that synovialcells have immune privilege and will not be rejected by a host organism[28,29]. Some studies supported the tolerance of allo- or xenotransplants fortissue healing [30,31]. An additional study showed that xenogeneic chondrocytescan successfully repair full-thickness cartilage defects [32]. However, in arecent 2010 study, xenotransplantation of porcine chondrocytes into rabbits wasunsuccessful for long term cartilage healing [11]. Our study provided a uniquecontribution to the relatively limited body of work in the literature usingthis type of co-culture model. Our data is in agreement with previousliterature suggesting that mesenchymal stromal cells maybe immunotolerated orimmunoprivileged [15, 17,33]. Findings in our human allogeneic experimentdemonstrated that no detectable immune activation was prompted; however, animmunosuppression was not observed as in other studies. In cell culture, MSCshave also been able to suppress T cell proliferation [13], sugCopyright © 2012SciRes. OJCB S. S. JUMP ET AL. 7 gesting MSC, including stem cells, are notonly immunotolerated, but may be immunosuppressive [15]. Maccario andcolleagues observed a decrease in the formation of CD8+ T cells and naturalkiller lymphocytes in favor of an expansion of CD4+ lymphocytes in response toallogeneic MSC [17]. However, in our xenogeneic experiments, the evidencesuggested that equine dermal fibroblasts may play a role in the inhibition ofan immune response. Our data showed less of an increase in CD8+ cells afterexposure to xenogeneic dermal fibroblasts. Additional evidence has shown thatfibroblasts, both autologous or allogeneic cells, resulted in no increase inT-cell proliferation; however, this effect required a modification of CD80expression on the surface of the cell [12]. Data is accumulating that suggestsan immunotolerance to selected allogeneic and xenogeneic isolated cells. Ourdata showed variability in human splenocyte initial cell proportions from humanto human, demonstrated as wide standard deviations in isolated hSpl numbers andpercentages of CD+ cells. The variability of splenocyte populations under ourconditions are likely explained by differences in age, gender, andhematological parameters of our human spleen donors. Human spleens wereacquired from an organ donor center, without exclusion for age, gender, orcause of death. Previous health history of the human spleen donors may havealtered the proportion of various cell types in the spleen including mature lymphocytes,T-helper cells, dendritic cells, or natural killer/ cytotoxic T cells. Theheterogeneous human population providing hSpl sources probably addedvariability to our co-culture condition responses, but represent a typicalrecipient population of humans. Despite the variability inherent in using hSplfrom multiple human donors, we were able to demonstrate the natural selectionand adaptation of hSpl to the cell culture environment in a shortterm cellculture study, minimal lymphocyte activation by allogeneic and xenogeneic cellexposure and the ability of donor allogeneic and xenogeneic cells to thrive indirect contact with a mixed population of immune cells. The clinicalapplicability of our work extends to potential cell-based strategies for the repairof human tissue. Allogeneic or xenogeneic cell sources could serve as apowerful clinical tool to enhance current medical practices that promotehealing. Finding a potential cell- or tissue-based treatment for cartilage isdependent upon further understanding of the immunoresponse of cells, includingsynovial lining cells or chondrocytes. Further characterizing theimmunoresponse may enable potential molecular or other cellular modificationsto enable specific cells to “tolerate” an immune response or modify the nativeimmune response. It is possible that specific cells could be engineered tomodify the acute immune response in vivo, ahead of or in conjunction cellularinjection or transplantation 5. Conclusions We uniquely investigated theimmunoresponse of a mixed population of hSpl to three different mechanisms ofallogeneic or xenogeneic challenge: 1) cellular antigens 2) cellular/adenoviralantigens and 3) lipopolysaccharide (LPS). In our in vitro model, neitherallogeneic nor xenogeneic cells elicited a lymphocyte/dendritic cellimmunoresponse. Our study provided evidence that, in a shortterm cultureenvironment, hSpl undergo a natural selection and adaptation to cell cultureconditions. In hSpl cocultures with hSD-MSC, hSD-MSC-GFP, eqDFb, or eqMSC, CD3+and CD8+ hSpl increased in proportion and decreased in number with time inculture, but not with exposure to allogeneic or xenogeneic cell antigens. Theevidence suggested that allogeneic or xenogeneic may be immunotolerated in theshort term and upon first exposure by human recipients. Further studies tocharacterize the long-term response in vivo and the anemnestic response iswarranted 6. Acknowledgements Dr. Jump was supported by a Fellowship at TheOhio State University from the Sports Medicine Center. Dr. Bertone wassupported by NIH/NIAMS grant number K08AR4920101. The work was supported by theTrueman Endowment. We would like to acknowledge and thank Dr. Akikazu Ishiharafor his assistance in the statistical and data analysis and for proofreadingand editing our mansucript. Additionally, we offer a special thanks to Dr.Prosper Boyaka and his laboratory personnel, Dr. Junbae Jee and Haley Steiner,for assistance with technical guidance and methodological expertise inperforming flow cytometry.